Intelligente celler, en billion ganger mer komplekse enn noen har forstått: Ved evolusjon, eller design?
Oversatt herfra.
Biolog Brian Ford:
 "Den mikroskopiske verdenen til den eneste, levende cellen speiler vår egen på så mange måter: celler er i det vesentlige autonome, følsomme og geniale. I livene til enkeltceller kan vi oppfatte røttene til vår egen intelligens. "
"Den mikroskopiske verdenen til den eneste, levende cellen speiler vår egen på så mange måter: celler er i det vesentlige autonome, følsomme og geniale. I livene til enkeltceller kan vi oppfatte røttene til vår egen intelligens. "
Det følgende er en imponerende, strålende beskrivelse av cellen som gir ære til vår ufattelige, uforståelige, mesterlige ingeniør, skaper, Gud og Herre.
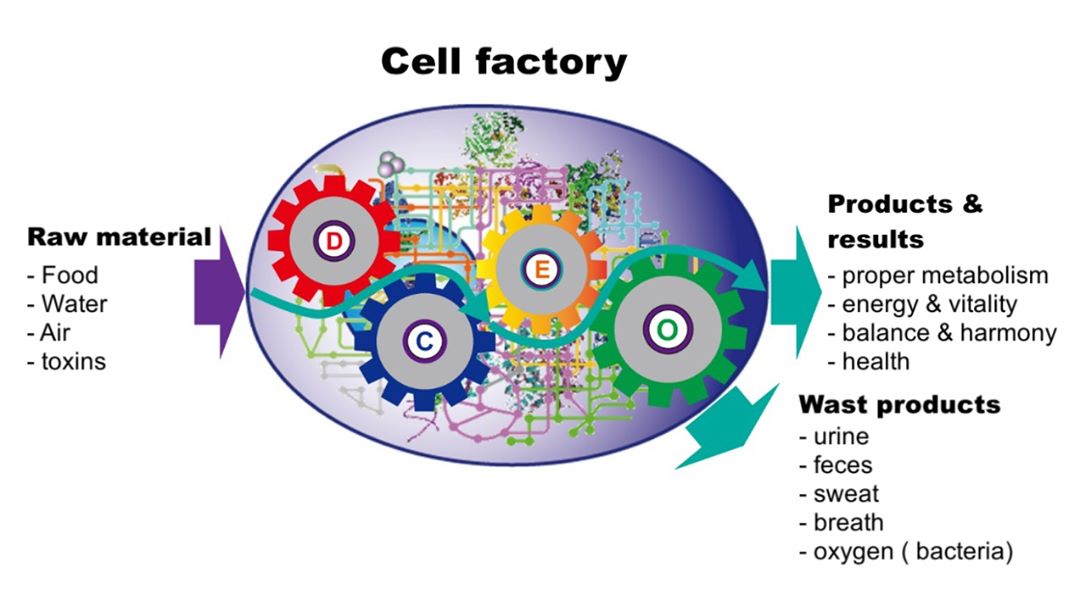
Når vi observerer atferd i en levende celle inkludert kontinuerlig og nøye koreografert bearbeiding av mitokondrier, endeløs migrering av granuler og tømming av vakuoler, ledning av diskrete partikler i toveis strømmer av cytoplasma som trafikk på en motorvei, de grundige endringene av kjernens posisjon i kiselalgene under deling, og den forsiktige inspeksjonen av byttedyr av et rovdyrsiliat, så kan vi tenke oss at cellen kan være en milliard, eller til og med en billion ganger mer kompleks enn noen har forstått. Vi har hevet blikket mer for å forstå omfanget av vår uvitenhet, og nå bør vi bruke vår egen innfødte intelligens til å prøve å løse opp fra hvor den kom fra. Hele den levende cellen er et uforståelig mirakel, og dens mangesidede evne til å kommunisere, ta beslutninger og svare på uforutsigbare situasjoner med en viss intelligens kan redegjøre for mange observerte fenomener som ellers er uforklarlige. Glem reduksjonisme: Hele den levende cellen, som en enhet, krever nå vår oppmerksomhet som aldri før.
Den levende cellen viser oppfinnsomhet, og kanskje til og med intelligens.
Hver celle vet sin passende størrelse, alder, tid på dagen og sin egen plassering. Minst tusen forskjellige typer nevroner har spesifikke størrelser og former for å passe inn i bestemte nevrale kretser. Nevroner avgir elektriske pulser med en frekvens på rundt 40 Hz, kjent som neuron pigger, hvis opptak er kjent innen nevrovitenskap. Timing av nevron-topper er nå anerkjent som viktig. Jeg ble fascinert av å se for meg piggene, ikke som raskt gjentatte signaler, men som diskrete impulser som hadde kodet informasjon.
 Det er mange forsøk på å etterligne hjernen gjennom digital modellering. For eksempel prøver Blue Brain-prosjekt3 i Sveits å skape en digital emulering av hele pattedyrshjernen ved å reversere hjernekretsene. Denne modelleringen av sinnet gjennom digitale data kan ikke lykkes, siden nevronet ikke er en enkel digital enhet. Et enkelt nevron ville være mer enn nok for oss å takle, og selv det er sannsynligvis ikke mottakelig for våre primitive metoder.
Det er mange forsøk på å etterligne hjernen gjennom digital modellering. For eksempel prøver Blue Brain-prosjekt3 i Sveits å skape en digital emulering av hele pattedyrshjernen ved å reversere hjernekretsene. Denne modelleringen av sinnet gjennom digitale data kan ikke lykkes, siden nevronet ikke er en enkel digital enhet. Et enkelt nevron ville være mer enn nok for oss å takle, og selv det er sannsynligvis ikke mottakelig for våre primitive metoder.
Å tro at generene alene formidler en celle, bare som en blåkopi, kan ikke støttes. Den levende cellen er villig og setter pris på dens individuelle evne til å analysere data og ta beslutninger. Den ufattelige kompleksiteten til en levende celle blir vanligvis ikke verdsatt; til og med en simulering av måten mitokondrier beveger seg på ville være utenfor våre nåværende konvensjoner. å emulere fragmenter av vår trivielle forståelse forklarer ikke deres intrikatitet, for celler er intelligente, og de lever liv som vi knapt har begynt å behandle.
Celler trenger å vite hvor de er av mange grunner. Men det er veldig utfordrende for et individ å vite sin nøyaktige beliggenhet i forhold til en stor omverden, uten GPS. Til tross for store vanskeligheter, kan enkeltceller bemerkelsesverdig gjøre komplekse beregninger og beslutninger basert på deres eksakte forhold til andre celler og organer.
Celler bruker data fra forskjellige kilder, for eksempel morfogene felt av diffusible molekyler og elektriske gradienter og komplekse nettverk av gener. I tillegg til å være i stand til å beregne mange forskjellige faktorer og ta hensyn til dem, signaliserer cellene samtidig frem og tilbake til andre celler, og deler informasjon som hjelper til med å bestemme den nøyaktige plasseringen.
Selv om det kan virke enkelt for celler å måle en gradient, er det faktisk ikke, fordi mange variabler kan påvirke målingen. For å finpusse nøyaktigheten av informasjon fra diffunderende molekyler, elektriske gradienter og mekaniske signaler, må den bruke tilbakemeldingsløkker, flere komparative målinger, tidsbestemte målinger, kompleks genregulering, prinsipper for selvorganisering og veldig spesifikk celle-til-celle-signalering. Celler blir ekte 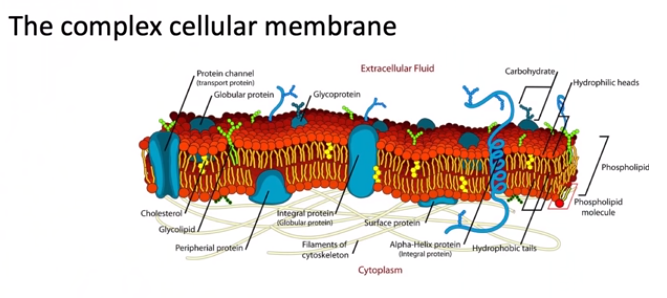 matematikere for å svare på spørsmålene. Dette innlegget vil beskrive noen av problemene som celler må overvinne for å ha nøyaktig stedsinformasjon som er kritisk for hjernestrukturer og immunfunksjon.
matematikere for å svare på spørsmålene. Dette innlegget vil beskrive noen av problemene som celler må overvinne for å ha nøyaktig stedsinformasjon som er kritisk for hjernestrukturer og immunfunksjon.
Celler er i stand til å analysere flere konkurrerende signaler og ta beslutninger om hvordan de skal brukes. Hvert signal utløser en kaskade som stimulerer spesifikke genetiske nettverk. Men noe av informasjonen er mer nøyaktig enn andre, og det må tas beslutninger for å bestemme den nøyaktige plasseringen.
Celler bruker stor kompleksitet i mobilspråket. En type kommunikasjon mellom celler er å berøre og direkte transportere data som med synapser. Overførte signaler inkluderer immuncytokiner og kjemokiner (tiltrekkingsmidler), nevrotransmittere av mange typer, små og store RNA, små peptider og hormoner. Spesielle plantesignaler er kinocidiner. Signaler kan fungere enten lokalt eller på veldig lang avstand. For at signalet skal fungere, må cellene produsere de spesifikke proteinreseptorene og ha dem klare i membranene.
Celler bruker molekyler som beveger seg mellom celler for mange lokale målinger. Hvordan kan en individuell celle være så intelligent? Hvordan kan en individuell celle integrere så mange forskjellige typer informasjon og deretter handle sammen med mange andre celler? Hvor ligger denne intelligensen? Hvor er retningen for alt dette? Cellen må være knyttet til andre informasjonskilder som for eksempel bevissthet.
---
Intelligens i celler: Evidens for intellgent Design
Oversatt herfra.
 Cellen er en organisme. Cellebiologi bør bli fokus for biovitenskap og den intelligente cellen finnes i dens kjerne. Handlingspotensialene som vi kan måle, kan sees på som et språk der nevroner overfører behandlede data fra en til den neste. Nevronene avgir et signal på ≈40 Hz, og på grunn av den akutte toppen av hver bølge i opptaket, blir disse signalene kategorisert som nevronpigger. Svarene fra celler er ikke mottagelige for prediktiv matematisk modellering. De er så forskjellige, komplekse og multivariate at de trosser rasjonell analyse.
Cellen er en organisme. Cellebiologi bør bli fokus for biovitenskap og den intelligente cellen finnes i dens kjerne. Handlingspotensialene som vi kan måle, kan sees på som et språk der nevroner overfører behandlede data fra en til den neste. Nevronene avgir et signal på ≈40 Hz, og på grunn av den akutte toppen av hver bølge i opptaket, blir disse signalene kategorisert som nevronpigger. Svarene fra celler er ikke mottagelige for prediktiv matematisk modellering. De er så forskjellige, komplekse og multivariate at de trosser rasjonell analyse.
Celler trenger å vite hvor de er av mange grunner. Men det er veldig utfordrende for et individ å vite sin nøyaktige beliggenhet i forhold til en stor omverden uten GPS. Til tross for store vanskeligheter, bemerkelsesverdig, kan enkeltceller gjøre komplekse beregninger og beslutninger basert på deres eksakte forhold til andre celler og organer.
Hvite blodlegemer på farten- T-celler, B-celler, makrofager og leukocytter - trenger å vite nøyaktig hvor en infeksjon er lokalisert og hvor de er i forhold til den. Nevroner i fosteret må finne sine nøyaktige steder i enormt komplekse nettverk. Celler som vokser i et organ eller en lem, må vite nøyaktig hvordan de passer inn i strukturen og formen og når de skal slutte å lage nye celler til slutt. Hvordan kan disse intelligente cellene vite nøyaktig sin plassering?
Ved å følge flere regler og målinger og syntetisere denne informasjonen, er celler i stand til å beregne hvor de er. Men det er mange mulige feilkilder som må tas i betraktning.
Celler bruker data fra forskjellige kilder, for eksempel morfogene felt av diffusible molekyler og elektriske gradienter og komplekse nettverk av gener. I tillegg til å være i stand til å beregne mange forskjellige faktorer og ta hensyn til dem, signaliserer cellene samtidig frem og tilbake til andre celler, og deler informasjon som hjelper til med å bestemme den nøyaktige plasseringen.
Selv om det kan virke enkelt for celler å måle en gradient, er det faktisk ikke, fordi mange variabler kan påvirke målingen. For å finpusse nøyaktigheten av informasjon fra diffunderende molekyler, elektriske gradienter og mekaniske signaler, må den bruke tilbakemeldingsløkker, flere komparative målinger, tidsbestemte målinger, kompleks genregulering, prinsipper for selvorganisering og veldig spesifikk celle-til-celle-signalering. Celler blir ekte matematikere for å svare på spørsmålene. Dette innlegget vil beskrive noen av problemene som celler må overvinne for å ha nøyaktig stedsinformasjon som er kritisk for hjernestrukturer og immunfunksjon.
Komplekse reise- og cytokinsignaler
Immunceller som beveger seg gjennom kroppen til et betennelsessted, må håndtere tusen forskjellige faktorer ved å bruke mange forskjellige mekanismer. I løpet av denne reisen må celler vite hvor de er og hvor de skal. Celler er avhengige av veldig detaljerte signaler, inkludert cytokiner, kjemokiner og nevrotransmittere, og gradienter fra mange forskjellige kilder. Celler holder også på kommunikasjonen mellom andre immunceller som farter.
Vevene skiller ut tiltrekkende molekyler som kan hjelpe dem gjennom blodkar, ekstracellulær matrise og organer. For denne typen oppførsel har immunceller mange forskjellige reseptorer som lar dem forstå flere signaler. Hvert kroppsvev - lymfe, lever, lunge, hud og hjerne - har også forskjellige stillaser som må krysses. Cellene navigerer ved hjelp av alle gradientene sammen med andre mekanismer.
Det produseres mange forskjellige gradienter som danner bølger av forskjellige typer informasjon. Celler er i stand til å analysere flere konkurrerende signaler og ta beslutninger om hvordan de skal brukes. Hvert signal utløser en kaskade som stimulerer spesifikke genetiske nettverk. Men noe av informasjonen er mer nøyaktig enn andre, og det må tas beslutninger for å bestemme den nøyaktige plasseringen.
Cellesignalering for nøyaktighet
Celler bruker stor kompleksitet i mobilspråket. En type kommunikasjon mellom celler er å berøre og direkte transportere data som med synapser. Overførte signaler inkluderer immuncytokiner og kjemokiner (tiltrekkingsmidler), nevrotransmittere av mange typer, små og store RNA, små peptider og hormoner. Spesielle plantesignaler er kinocidiner. Signaler kan fungere enten lokalt eller på veldig lang avstand. For at signalet skal fungere, må cellene produsere de spesifikke proteinreseptorene og ha dem klare i membranene.
Små nanorør, bygget med aktinmolekyler omgitt av membraner, finnes mellom nesten alle celler. Faktisk er disse cytonemene den viktigste måten virus overføres mellom celler på. Cytonemer brukes også til horisontal genoverføring.
 En annen universell teknikk for mobilkommunikasjon er vesikler. Eksosomer utskilles fra membranene i celler og tas deretter inn av andre celler (som endosomer). Det er nå kjent at nevroner og glia kommuniserer med disse vesiklene og sender viktig protein og genetisk informasjon inkludert kritiske RNA.
En annen universell teknikk for mobilkommunikasjon er vesikler. Eksosomer utskilles fra membranene i celler og tas deretter inn av andre celler (som endosomer). Det er nå kjent at nevroner og glia kommuniserer med disse vesiklene og sender viktig protein og genetisk informasjon inkludert kritiske RNA.
Enda mer uvanlig er de mange forskjellige måtene cellene bruker elektrisitet og elektriske gradienter på. I hver av disse kommunikasjonstypene er cellen på jakt etter målenøyaktighet. De kan sammenligne disse signalene med forskjellige typer informasjon for å bestemme nøyaktigheten.
Mobilmålinger av plassering - behovet for nøyaktighet
Målinger i den menneskelige verden gjøres gjennom målebånd for korte avstander og radiobølger for lange avstander. Celler bruker molekyler som beveger seg mellom celler for mange lokale målinger. Disse molekylene overholder de enkle lovene for diffusjon i en vannløsning. Men i motsetning til radiobølger kan diffusjon oppstå i forskjellige hastigheter, og bevege seg saktere når den sprer seg fra kilden. Når molekylene danner en stabil gradient (en stabil tilstand), kan cellen forstå hvor de er i gradientfeltet. Men mange graderinger er ikke stabile. Så cellen må iverksette forskjellige metoder for å sikre at målingen deres er nøyaktig.
Mange faktorer gjør gradientinformasjon upålitelig. Molekyler produseres i forskjellige hastigheter, og reseptorer varierer når det gjelder å aktivere signalmolekyler i deres konsentrasjoner. De diffunderbare molekylene elimineres på forskjellige måter. Den måten en reseptor fungerer på, kan påvirke nøyaktigheten. Miljøfaktorer som temperatur eller hastighetsbegrensende næringsstoffer påvirker nøyaktigheten.
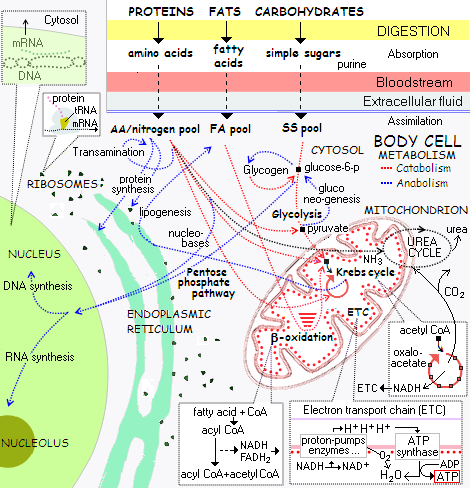 Andre variabler er regulering av trinn, tatt for å produsere og eliminere et molekyl som brukes for gradienten. Det diffusible molekylet kan fremstilles av cellene, eller metaboliseres fra andre molekyler i flere trinn.
Andre variabler er regulering av trinn, tatt for å produsere og eliminere et molekyl som brukes for gradienten. Det diffusible molekylet kan fremstilles av cellene, eller metaboliseres fra andre molekyler i flere trinn.
Et eksempel forekommer i embryoet, der en gradient er retinsyre. Retinsyre danner mønsteret som den bakre delen av hjernen er bygget på. Retinsyre er laget i to metabolske trinn fra inntatt vitamin A. Denne enkle metabolske veien er sterkt regulert for å opprettholde gradienten. å opprettholde gradienten innebærer tilbakemeldings-løkker til molekylene i sin syntese, metabolsk nedbrytning og reseptormekanismene i cellene. Spesielle proteiner er involvert i regulering av retinsyre, det samme er cytokrom P450-enzymer som eliminerer den. Når eksperimenter endrer mengden, opprettholder disse kontrollveiene gradienten og derfor informasjonen nevronene bruker for å bygge hjernevevet.
Ett sett med feil oppstår når miljøfaktorer påvirker alle celler på samme måte, for eksempel temperatur. Andre feil oppstår når hver celle reagerer forskjellig på en miljøfaktor.
Noen celler kan i seg selv endre hvordan gradienten produseres. En celle kan også øke eliminasjonen av gradientmolekylet. Det kan prøve å stabilisere signalet ved å produsere flere reseptorer. Men disse manøvrene produserer andre problemer ved å ha en metning av reseptorer og ytterligere unøyaktighet. Cellen kan måle ettersom gradienten utvikler seg, men dette må tidsbestemmes nøyaktig. Celler kan også introdusere et annet molekyl som samhandler med det diffuserende signalet.
En annen måte celler forbedrer nøyaktigheten på er å unngå å måle diffusjon og i stedet bruke aktiv transport. I planter er det ingen steder å diffundere mellom celler, og derfor er aktiv transport en kritisk faktor for å bestemme hvor celler er. Vannløselige molekyler transporteres aktivt over plantemembraner.
To andre situasjoner skaper forskjellige måleproblemer. Mange små celler produserer større svingninger i måling. Noen få store celler med et fast større antall reseptorer har problemer med metning.
Generelt sett blir andre verre ved å forbedre nøyaktigheten til en hvilken som helst faktor. For å bruke graderinger, må spesifikke faktorer vektlegges som er mest pålitelige for sammenligning. Når gradienten er etablert i stabil tilstand, inkluderer noen pålitelige faktorer som ikke endrer diffusjon temperatur, metabolske celletilstander, hindringer og klebrig gjenstander. Cellen kan ta hensyn til disse på forskjellige måter for å fastsette målingene.
Mengdemål
Oppsummert, kan celler utføre flere målinger, via det omkringliggende elektromagnetiske feltet og deretter syntetisere disse til bruk for posisjonsinformasjon - enten den samme faktoren over tid eller flere faktorer. Hver av beregningene må være uavhengige av de andre. I den bakre hjernen bruker foster-stamcellene ikke bare retinsyre for gradienten, men bruker også to andre molekyler på samme tid - FGF og Wnt. Mange andre celler bruker flere forskjellige diffusible gradienter.
En teknikk for å samle flere målinger over tid er å hente signalet og plassere det i et endosom. Inne i cellen i en vesikkel kan signalet fortsette å måles sammen med andre signaler.
En annen mekanisme for telling er å ha en lengre eller kortere virkning ved reseptorkomplekser. Når flere signaler samhandler med komplekset før det reagerer, kan et gjennomsnitt måles. Faktorer som forstyrrer gradienter i kort tid, vil bare påvirke noen av signalene, og gjennomsnittet vil ikke være basert på kortvarig forstyrrelse.
Men forstyrrelser som varer lenge trenger forskjellige mekanismer. Dette kan oppstå når det er store forskjeller i antall cellereseptorer, hvis produksjon tar lang tid. I denne situasjonen, hvis cellen må ta avgjørelsen raskt, vil ikke informasjonen fungere, men hvis den trenger lengre tid, kan informasjonen være nøyaktig. Bruk av tidsbestemte gjennomsnitt innebærer også at gradientmolekylet har kort halveringstid. Cellene må vente lenger på å ta sin beslutning.
 Noen celler bruker en bryter. Når den er slått på, via en viss mengde signalering, forblir den på til det er en stor endring, til å snu den av. Dette skaper en tidsperiode som signalet er på.
Noen celler bruker en bryter. Når den er slått på, via en viss mengde signalering, forblir den på til det er en stor endring, til å snu den av. Dette skaper en tidsperiode som signalet er på.
Under fosterutvikling må stamceller ofte bestemme om de vil dele seg eller ikke på et bestemt sted. Disse typer beslutninger tas av en brytermekanisme som har forskjellige terskler. Mange av disse bryterne påvirker endringen i genetiske nettverk. Disse er avhengig av mengden svingninger som oppstår (støy).
En annen teknikk er å måle forholdet mellom reseptorer som utløses sammenlignet med dem som ikke er det (dette gjøres med Hedgehog).
Bruk av andre celler for måling
Bakben av en honning-bi celler som er nær hverandre, kan legge informasjonen sammen. Individuelle celler foretar en måling og signaliserer deretter til naboceller om innvirkning på komplekser inne i kjernen. Et eksempel oppstår når du danner en vinge fra et gradientsignal. Signalet utløser en transkripsjonsfaktor. Ved kanten av formingsfløyen er det veldig lave signaler som er unøyaktige og som ikke utløser bryteren. I stedet kommuniserer cellene med hverandre slik at celler bare blir utløst når de ligger rett ved siden av en celle som allerede er utløst. Dette skjer også gjennom tidsbrytere. Med disse multiple signalene blir vingen bygd ut fra sentrum til kanten, hvor den stopper på et nøyaktig punkt.
Andre problemer og løsninger
Hvite blodlegemer: Celler er ikke alltid i stand til å foreta uavhengige målinger av flere faktorer. Alle faktorene kan bli påvirket av en bestemt variabel. Hvis varme øker tettheten til gradienten, kan cellen måle et annet molekyl som påvirkes forskjellig av varmen og sammenligne dem. Disse signalene kan da avbryte hverandre. Denne typen måling skjer når visse faktorer har kjent påvirkninger som endrer flere gradientmolekyler som måles. Hvis en miljøendring kan gi flere forskjellige effekter som er kjente, kan forskjeller beregnes for å bestemme plasseringen.
 Et eksempel på bruk av to signaler oppstår ved endring av diffusjonshastigheten til signalmolekylet og samtidig en endring i antall reseptorer i cellen som tar opp signalet. Disse to endringene påvirker hverandre, og cellen kan gjøre en kompleks bestemmelse ut fra disse målingene for plasseringen. På en eller annen måte er celler i stand til å måle disse matematiske abstraksjonene. I vingeeksemplet gitt ovenfor reagerer signalet på forskjellene mellom celler i forhold til gradienten.
Et eksempel på bruk av to signaler oppstår ved endring av diffusjonshastigheten til signalmolekylet og samtidig en endring i antall reseptorer i cellen som tar opp signalet. Disse to endringene påvirker hverandre, og cellen kan gjøre en kompleks bestemmelse ut fra disse målingene for plasseringen. På en eller annen måte er celler i stand til å måle disse matematiske abstraksjonene. I vingeeksemplet gitt ovenfor reagerer signalet på forskjellene mellom celler i forhold til gradienten.
Noen brytere tar hensyn til flere signaler over en bestemt tid, men endrer svarene basert på hvor mye tilfeldig bakgrunnsstøy det er. Forståelse av mengden tilfeldig signalering i forhold til det virkelige signalet kan beregnes.
Selvorganiserende strukturer blant flere celler
Noen signaler forekommer bare mellom lokale celler. Men signalet kan være kraftig og spre seg fra en lokal gruppe til en annen som en bevegelig front. Studier viser at det er en grense for hvor langt denne typen informasjon kan spre seg, og dermed lage en struktur. Andre tilbakevendende gradienter kan da forekomme. Effekten av disse spredningsbølgene kan også måles og sammenlignes med en stabil bølge.
Et eksempel på denne spredte bølgen av informasjon og signalisering i menneskelivet oppstår når vi bruker lysets hastighet og sammenligner den med tordenhastigheten for å bestemme hvor langt lynet skjedde. Celler gjør en lignende beregning.
Flere forskjellige interaksjoner mellom lokale celler kan skape romlig orden. Celler som er en del av et innledende signal, for eksempel Notch-banen, kan også delta i en annen type signalering. Disse sekundære gradientene organiserer flere celler på forskjellige måter. Én type sekundært signal kan hemme det originale signalet ved slutten av organet. Ulike nivåer av celleadhesjon kan være en del av denne signaliseringen. å sette alle disse sammen kan skape veldig organiserte mønstre som kreves i organer og fosterhjernen.
Andre stedsignaler kan også kobles til disse selvorganiserende systemene. Selvorganiserende signaler kan også være korrelert med et bestemt gradientsignal som noterer den nøyaktige plasseringen der de skal organiseres. Organiseringen av organet og posisjonsfølelsen til de enkelte cellene kan være en del av denne samme større prosessen.
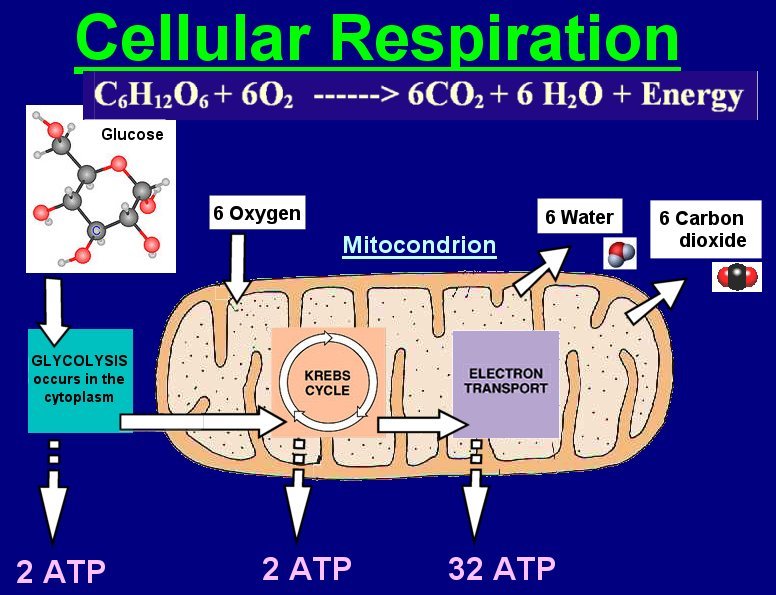 Intelligente celler kjenner sin plass
Intelligente celler kjenner sin plass
Hentet herfra.
Celler trenger å vite nøyaktig hvor de er av mange grunner - for å bygge fosterets hjerne, reise til et infeksjonssted og opprettholde kroppsorganer. De bruker mange forskjellige typer informasjon for å svare på dette overraskende komplekse spørsmålet og må syntetisere dem alle for å ta beslutninger. Problemer oppstår med nøyaktigheten av informasjonen, for eksempel å bruke gradienter laget av diffuserende molekyler eller elektriske gradienter. Hver situasjon kan involvere mange forskjellige variabler som kan endres med temperatur, metabolisme og andre miljøfaktorer. Hver celle må kunne bruke avansert matematikk for disse bestemmelsene, og utvinne stabil informasjon fra flux.
Andre innlegg har beskrevet det komplekse språket for cellulær signalering ved hjelp av cytokiner, kjemokiner, nevrotransmittere, elektriske signaler, foton-signaler, nanorørssignaler og vesikler fylt med genetisk informasjon. Celler bruker alle disse kommunikasjonsmetodene for å sammenligne informasjon med lokale celler. Nærliggende celler kan starte en ny gradient for å stoppe prosessen og markere et endepunkt for veksten av et organ eller lem.
Hvordan kan en individuell celle være så intelligent? Hvordan kan en individuell celle integrere så mange forskjellige typer informasjon og deretter samhandle med mange andre celler? Hvor ligger denne intelligensen? Hvor er retningen for alt dette? Cellen må være knyttet til andre informasjonskilder som for eksempel bevissthet.
Oversettelse og bilder ved Asbjørn E. Lund